От одной клетки к множеству
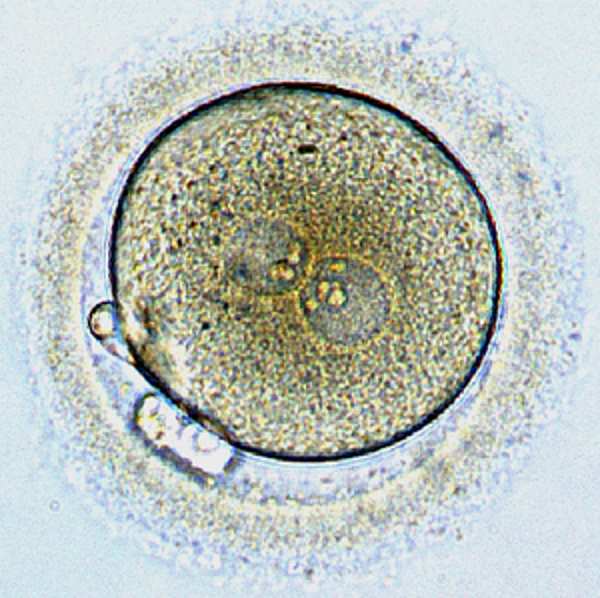
Зигота - оплодотворенная яйцеклетка, одноклеточная стадия развития эмбриона
Величайшая ирония биологии в том, что человеческий организм – одна из самых сложных сущностей во Вселенной, – имеет очень простое начало. Взрослый человек состоит из миллионов миллионов клеток. Их в десять раз больше, чем звезд в нашей галактике, или, если привести более земное сравнение, в десять раз больше, чем песчинок на небольшом пляже. Все это огромное количество клеток не свалено как попало, а организовано в сложные структуры, и, несмотря на многовековую историю анатомических исследований, мы все еще не знаем всех деталей их строения. Существуют сотни типов клеток, и у каждого своя функция и свой образ жизни, при этом клетки каждого конкретного типа появляются и обновляются в нужном количестве и в нужном месте. Вся эта сложнейшая конструкция развивается из одной оплодотворенной яйцеклетки. И скромной клетки, внешне лишенной каких-либо особых признаков. И уже с самого начала постепенное усложнение человеческого организма протекает в режиме самоорганизации: ему приходится действовать самостоятельно, без какой-либо посторонней помощи.
Первый шаг к усложнению – переход от одной клетки к множеству. Это обусловлено необходимостью одновременного протекания большого количества различных процессов. Например, сейчас вы дышите, перевариваете пищу и выводите из организма вредные вещества; ваши волосы растут, кожа обновляется, кровь фильтруется, иммунная система борется с болезнетворными бактериями, температура тела регулируется; вы слушаете, читаете и думаете, а теперь, когда вы дочитали это предложение до конца, возможно, также прислушиваетесь к своему организму. Эти процессы, как и многие другие, приводятся в действие с помощью особых белков и биохимических механизмов. Многие из этих процессов никак не могут протекать в одном и том же месте. Например, представим, что в одном и том же месте организма матери вырабатывается молоко для ребенка и тут же переваривается то молоко, которое она только что выпила с чаем. Есть и другие примеры процессов, несовместимых по более сложным причинам: из-за особенностей строения белков или функционирования генов.
Сложные организмы справляются с этой проблемой благодаря компартментализации – принципу разделения процессов в пространстве. Тело состоит из органов, имеющих определенные функции, органы – из специализированных тканей, а ткани – из разных типов клеток. Однако внутри клетки большая часть молекул постоянно находится в движении, из-за чего трудно достичь одновременного выполнения многих операций. Компартментализация существует и на уровне клетки. Однако «многозадачность» клетки имеет свои пределы. Поэтому мы будем считать клетку базовой единицей, которая одновременно выполняет лишь одно-два дела. Именно поэтому многообразие типов клеток является необходимым шагом к созданию сложного организма.
Механизмы, за счет которых одна клетка превращается в две, а потом и во множество клеток, не только принципиально важны для эмбрионального развития, но и ярко демонстрируют возможности самоорганизации. Простые маленькие молекулы могут самоорганизовываться в крайне сложные структуры, имеющие гораздо больший пространственный масштаб, чем сами молекулы, причем без какого бы то ни было предварительного плана. Это краеугольный камень, лежащий в основе понимания развития эмбриона. Поэтому, мы подробно остановимся на механизмах деления клетки, а в дальнейшем будем принимать их как данность.
Оплодотворенная яйцеклетка, с которой начинается развитие человека, необычно велика. Она достигает десятой доли миллиметра в диаметре и видна даже невооруженным глазом. Большинство клеток организма намного меньше: примерно сотая доля миллиметра в диаметре и тысячная доля объема яйцеклетки. Это означает, что оплодотворенная яйцеклетка может превратиться в многоклеточный эмбрион просто поделившись на две, затем на четыре, на восемь и так далее, без перерывов на рост. Такой тип деления клетки – дробление – очень удобен для эмбриона, так как позволяет отложить проблему питания, обеспечивающего энергию для роста, на потом, а именно на этап, когда эмбрион уже станет многоклеточным и сможет выделить для переработки пищи специализированные части тела.
Если расти не нужно, процесс деления сводится к распределению молекул (например, белков) поровну между дочерними клетками. Суть деления при неизменном объеме заключается в сохранении концентрации белков и питательных веществ. Ярким исключением из этого правила является молекула ДНК: в исходной клетке сорок шесть хромосом (двадцать три от матери и двадцать три от отца), и каждая новая клетка должна содержать такое же их число. Поэтому хромосомы должны копироваться (реплицироваться) перед каждым делением клетки. Более того, должны существовать специальные системы, гарантирующие «честное» распределение реплицированных хромосом по дочерним клеткам – каждая из них должна получить не сорок шесть любых хромосом, а по одной копии каждой хромосомы от отца и по одной – от матери. Система, которая обеспечивает эту нелегкую задачу, является одной из основных систем клеток животных и растений и существует уже около 2,5 млрд лет. И всего пару миллионов лет назад появились существа, которые в принципе способны понять, как она работает.
Копирование ДНК – самая простая и самая древняя часть процесса, ей уже как минимум 3,5 млрд лет. Она основана на том, что молекулы ДНК существуют в виде пары нуклеотидных цепей (иногда их называют «нитями»). Аденину на одной цепи всегда соответствует тимин на другой цепи, а цитозину – гуанин. Это строгое правило, связанное с химической структурой нуклеотидов, означает, что каждая цепочка содержит всю необходимую информацию о последовательности соседней цепи. Репликация ДНК начинается с того, что ферментный комплекс отделяет две материнских цепи друг от друга. Затем он собирает новую цепь для каждой из них, соединяя нуклеотиды в порядке исходной цепи. Каждая новая цепь соединяется со старой, которая служила для нее матрицей. В результате получаются две молекулы ДНК вместо одной. То есть происходит репликация ДНК. Белки, в которые завернута ДНК, добавляются сразу же после копирования.
После репликации сорок шесть хромосом одноклеточного зародыша должны быть распределены так, чтобы каждой дочерней клетке непременно досталось по одной копии каждой материнской хромосомы и каждой отцовской хромосомы. Этот процесс можно разбить на несколько этапов:
1) определение центров двух дочерних клеток;
2) выравнивание всех скопированных хромосом между этими центрами;
3) «растаскивание» копий – по одному экземпляру каждой пары отходит каждой дочерней клетке;
4) отделение дочерних клеток друг от друга. Каждый из этих этапов включает в себя координированные действия в пространственном масштабе, значительно превышающем размеры вовлеченных в них молекул.
При этом весь процесс должен протекать без отклонений, несмотря на то что точное расположение основных компонентов (хромосом, например) будет постоянно меняться. Поэтому все эти этапы в значительной степени полагаются на адаптивную самоорганизацию и могут служить отличным примером для иллюстрации этого принципа.
Первая проблема заключается в выявлении центров новых дочерних клеток. Проще всего понять, как определяется центр в типичной взрослой клетке, которая не собирается делиться, а просто находится в состоянии покоя. На первый взгляд проблема кажется простой. Однако при более детальном рассмотрении все усложняется. Клетки вообще не имеют четкой формы: она зависит от их окружения. Это исключает любой заранее заготовленный план. Диаметр типичной клетки человека – примерно одна сотая миллиметра – кажется нам небольшим, ведь мы состоим из миллионов клеток. Однако это в тысячу раз больше, чем длина обычной молекулы белка. Тем не менее комплексы белков каким-то образом находят центр клетки. Это все равно что запустить в Альберт-Холл глухих людей с завязанными глазами и попросить их найти его середину.
Клетка нашла крайне остроумный способ решения этой проблемы. Он хорошо иллюстрирует, насколько важны могут быть тривиальные детали биохимических процессов для функционирования клетки. «Звездой» всего действа является белок тубулин, молекулы которого связываются друг с другом и образуют длинные структуры – микротрубочки. Одна из особенностей сборки молекул тубулина заключается в том, что объединение нескольких молекул тубулина для образования новой микротрубочки – событие маловероятное, а процесс присоединения молекулы тубулина к уже существующей микротрубочке, то есть ее удлинение, протекает относительно легко. Поэтому микротрубочки, как правило, не образуются спонтанно, но после того, как они образовались, они способны к спонтанному росту.
Вторая особенность биохимии тубулина заключается в том, что каждая отдельно взятая молекула может находиться в одном из двух состояний, «свежем» или «несвежем». «Свежие» молекулы медленно превращаются в «несвежие». Только «свежие» молекулы могут присоединяться к концам существующих микротрубочек. Концы микротрубочек устойчивы только тогда, когда они созданы из «свежего» тубулина (если концы остаются «свежими», то не имеет значения, утратит ли тубулин «свежесть» по всей длине микротрубочки). Если на конце микротрубочки «несвежий» тубулин, то конец начинает распадаться, и распад продолжается до тех пор, пока микротрубочка не присоединит стабильный «свежий» тубулин. Учитывая, что тубулин вдали от концов микротрубочки, скорее всего, был в ее составе дольше, чем тот, из которого состоит ее конец, эти «внутренние» молекулы, скорее всего, давно испортились, и «свежего» тубулина, способного предотвратить разрушение микротрубочки, не осталось. В таком случае микротрубочка распадется. Единственный способ избежать распада, не прибегая к помощи других молекул, – это быстрый рост, при котором «свежий» тубулин присоединяется к концу микротрубочки быстрее, чем разрушается «несвежий». Таким образом, микротрубочки либо быстро растут, либо катастрофически быстро распадаются. Существует постоянная вероятность разрушения, а это означает, что длинных трубочек всегда меньше, чем коротких. Эта особенность имеет непосредственное отношение к механизму нахождения клеточных центров.
Молекулы тубулина редко самопроизвольно объединяются в новые микротрубочки, и поэтому в клетке есть особые комплексы белков, которые могут катализировать этот процесс. Эти комплексы располагаются в ключевом месте клетки, а именно в центросоме, от которой микротрубочки расходятся радиально, как спицы от ступицы колеса. Пока они растут достаточно быстро для того, чтобы тубулин на их концах оставался свежим, микротрубочки будут удлиняться по направлению к периферии клетки. Существует две теории о том, как они помогают центросоме попасть в центр клетки. Они основаны на экспериментальных данных, полученных при исследовании различных организмов. Еще не ясно, какая из них справедлива для эмбрионов человека; не исключено, что обе. Одна из теорий связана с отталкиванием, а другая – с подтягиванием.
Механизм отталкивания основан на способности растущих микротрубочек отталкиваться от внутренней поверхности клеточной мембраны. Если центросома располагается близко к одной из сторон клетки, даже короткие микротрубочки смогут достичь поверхности мембраны и оттолкнуться от нее. В результате центросома отдаляется от этой стороны. Противоположной стороны клетки достигают только очень длинные микротрубочки, но они по упомянутым выше причинам встречаются редко. А поскольку таких микротрубочек меньше, они будут слабее отталкивать центросому от этой стороны клетки. Такое неравномерное распределение сил оттолкнет центросому от ближайшей мембраны, и она займет стабильное положение только тогда, когда силы отталкивания придут в равновесие. Равновесие же наступает тогда, когда центросома находится на одинаковом расстоянии от всех сторон: другими словами, в центре клетки (рис. 1, а). Ученые поместили центросомы в искусственно изготовленную «ячейку» и доказали, что им удалось найти центр «ячейки» с помощью механизма отталкивания.
Механизм подтягивания основан на действии небольших моторных белков, распределенных по всей клетке. Они могут связываться с микротрубочками и перемещаться по ним в сторону центросомы. Продвигаясь к центросоме, каждый из этих белков генерирует силу, которая слегка тянет микротрубочку в противоположном направлении, смещая ее в сторону клеточной мембраны. Так человек, идущий вперед по лодке, толкает ее, смещая назад. Чем длиннее микротрубочка, тем больше моторных белков могут связаться с ней и тем сильнее они за нее тянут в нужном направлении. Таким образом, если центросома ближе к одной стенке клетки, чем к другой, моторные белки сильнее всего тянут за длинные микротрубочки, направленные к дальней стороне, смещая центросому к центру клетки (рис. 1, б). Тщательные исследования оплодотворенных яйцеклеток просто устроенных организмов (таких, как морской еж или нематода) показали, что важную роль в этих клетках играет именно механизм подтягивания. Например, если часть микротрубочек разрезать с помощью лазера, центросома отскочит назад так, словно бы ее удерживали на месте натянутые микротрубочких. Не исключено, что в некоторых клетках работают оба механизма: сильное натяжение длинных микротрубочек «кнаружи» еще больше снижает их способности толкать центросому и усиливает дисбаланс сил отталкивания.
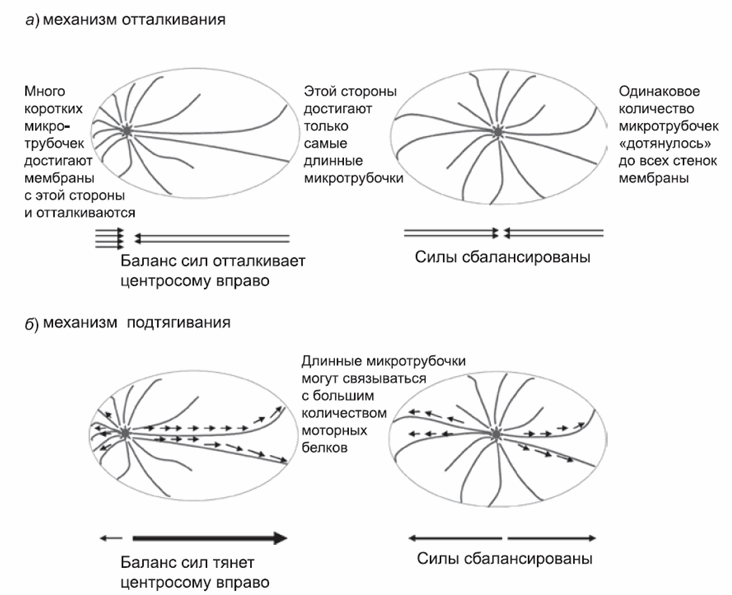
Рис. 1. Две теории, объясняющие, как центросома находит центр клетки с помощью микротрубочек. В модели отталкивания (а) микротрубочки отталкиваются от клеточной мембраны. Коротких трубочек всегда больше, чем длинных. Значит, центросома отталкивается сильнее от ближайшей клеточной мембраны. В модели подтягивания (б) моторные белки, распределенные по всей клетке, прикрепляются к длинным микротрубочкам. Коротких микротрубочек много со всех сторон центросомы, а длинные могут сформироваться только с удаленной от мембраны стороны. Поэтому центросому тянут к дальней стороне клетки и она удаляется от ближайшей мембраны
Однако какой бы из этих механизмов ни использовался клетками эмбриона человека, результат будет одним и тем же: центросома отцентрирует себя автоматически, хотя ни один из задействованных в этом процессе компонентов клетки не «знает» ее формы и не руководствуется для поиска центра какой-либо системой координат. Система организует себя сама. За такую самоорганизующуюся систему, способную адаптироваться практически к любым условиям, приходится расплачиваться энергией, необходимой для сборки микротрубочек и работы моторных белков; высокие энергетические затраты вообще характерны для адаптивной самоорганизации.
При клеточном делении необходимо определить не просто центр одной клетки, а те точки, которые будут центрами двух дочерних клеток, чтобы хромосомы переместились в правильном направлении. К счастью, клетка может при помощи того же механизма определить и два центра; все, что для этого нужно, – это две центросомы.
Центросомы состоят из «облака» белков вокруг пары соединенных вместе трубчатых структур, содержащих тубулин. Эта пара является организационным центром для материала центросомы. В процессе подготовки к делению клетки трубочки отсоединяются друг от друга, и каждая из них сразу же становится матрицей для сборки недостающего партнера. Таким образом, через некоторое время по соседству будут располагаться уже две пары трубчатых структур. Каждая из них организует вокруг себя центросомный материал и инициирует образование новых микротрубочек, радиально расходящихся от центросомы. В клетке с двумя центросомами радиальные микротрубочки одной системы «наталкиваются» на микротрубочки другой. В модели отталкивания микротрубочки одной системы будут отталкиваться от микротрубочек другой системы, точно так же как от клеточной мембраны. Присутствие второй центросомы и второй системы микротрубочек создает «ложное впечатление» о том, насколько близко каждая из центросом находится к клеточной мембране. Поэтому каждая из центросом оказывается не в центре клетки, а на максимальном удалении от другой центросомы (рис. 2). Аналогичным образом, в модели подтягивания с каждая система, состоящая из центросомы и микротрубочек, служит щитом для другой и не дает утащить центросому к дальней стороне клетки. Оба механизма, которые в клетках человека могут работать одновременно, будут иметь один и тот же эффект: ни одна центросома не будет находиться в центре клетки. Вместо этого они займут положение примерно посередине между истинным центром и периферией клетки (рис. 2). Таким образом, две центросомы определяют будущие центры двух новых клеток, которые образуются при делении материнской клетки. Опять же, это происходит «автоматически» – участники процесса ничего не «знают» о форме клетки.
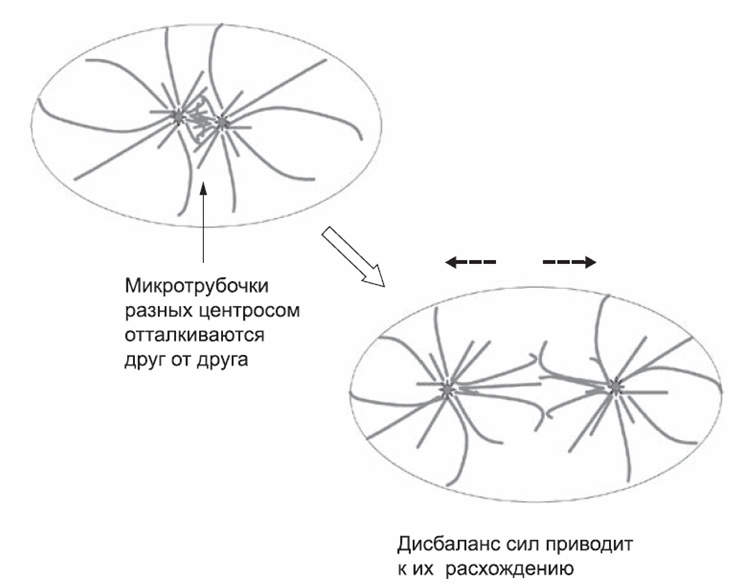
Рис. 2. Когда в клетке две центросомы, взаимодействие микротрубочек приводит к их расхождению
Радиальные микротрубочки, отходящие от центросом, служат не только для определения будущих клеточных центров. Они также способствуют расхождению двойного набора хромосом, чтобы каждая новая клетка получила их полный комплект. Чтобы это произошло, микротрубочкам сначала нужно соединиться с хромосомами. Опять же, ни одному из компонентов не требуется «знание» о положении других участников процесса. Система снова пользуется нестабильностью микротрубочек, которые проходят периоды сборки и катастрофического распада. Незащищенные концы микротрубочек особенно подвержены разборке, но, если они будут окружены особыми белками, способными связываться с микротрубочками, ситуация немного стабилизируется. В каждой хромосоме есть один специальный участок, содержащий такие белки, поэтому любая растущая микротрубочка может случайно «наткнуться» на этот участок и получить защиту. Таким образом, система, в которой микротрубочки расходятся случайным образом и не погибают только в том случае, если сталкиваются с хромосомой, приводит к тому, что все хромосомы оказываются «заякорены» при помощи микротрубочек.
Простейшей связи микротрубочек с хромосомами достаточно, чтобы каждая хромосома переместилась к центрам дочерних клеток, но для деления клетки требуется нечто большее. Одна копия хромосомы, скажем 9-й хромосомы отца, должна присоединиться к микротрубочке одной из центросом, а вторая ее копия должна присоединиться к микротрубочкам другой центросомы. Таким образом, каждая из них направится в свою клетку. Это достигается еще одним воздействием на устойчивость микротрубочек. Две копии каждой хромосомы, образовавшиеся в процессе репликации ДНК, соединены с помощью специальных белковых комплексов. Эти комплексы находятся в состоянии механического напряжения, когда два набора микротрубочек и связанные с ними моторные белки играют в «перетягивание каната» и пытаются растащить две копии хромосомы к противоположным центросомам. В этой ситуации генерируется сигнал, который приводит к более надежной стабилизации микротрубочек, чем в отсутствие механического напряжения. Если обе копии хромосомы прикреплены к микротрубочкам из одной системы, напряжения не возникнет, и микротрубочки долго не продержатся. Если они прикреплены к микротрубочкам разных центросом, которые пытаются оттащить их друг от друга, сигнал стабилизации достаточно сильный, и у микротрубочек гораздо больше шансов на выживание. Поэтому система постоянно меняется, продолжает находиться в режиме «поискового поведения», пока не находит то самое состояние, в котором копии хромосом будут растаскиваться в противоположных направлениях. Этот способ требует больших энергетических затрат, но зато действует автоматически и позволяет справиться даже с дополнительными хромосомами, появившимися в клетке в процессе эволюционных изменений или в ходе экспериментов.
Когда все хромосомы выстроились в линию, клетка может перейти к следующему этапу деления, на котором белки, связывавшие сестринские копии хромосомы, освобождают их, позволяя смещаться к противоположным сторонам клетки. Процесс не начнется до тех пор, пока все хромосомы не выстроены правильно, иначе дочерние клетки получат неправильное число хромосом и лишатся важных генов. Поэтому должна существовать система, предотвращающая преждевременное расхождение. В очередной раз используется способность белков, соединяющих копии хромосом, «ощущать» напряжение, возникающее, когда копии хромосомы крепятся к микротрубочкам из двух разных систем. Когда напряжение отсутствует, белковые комплексы продолжают генерировать сигнал, небольшую молекулу, которая может распространиться по клетке и блокировать переход к следующей стадии клеточного деления. Она как будто кричит «Не сейчас!» на языке биохимии. До тех пор пока все пары хромосомы не будут прикреплены к правильным микротрубочкам и не будут подвергаться растаскиванию, сигналы будут продолжаться и клетка будет ждать. Только тогда, когда натяжение всех белковых комплексов будет отрегулировано, сигналы смолкнут и можно будет начинать следующий этап деления. Опять же, эта система может приспособиться под любое количество хромосом.
Когда все хромосомы выстроились в середине веретена деления, а сигналы «Не сейчас!» затихли, клетка готова к следующему этапу. Белковые комплексы, которые соединяют пары копий хромосом, отпускают их, позволяя моторным белкам свободно подтягивать хромосомы по микротрубочкам к соответствующим центросомам. После того как все хромосомы распределились, еще одна автоматическая система размещает кольцо сократительных белков по «экватору» клетки между полюсами, положение которых определяют центросомы. Эти белки могут скользить относительно друг друга, образуя затягивающийся «пояс» клетки. Этот «пояс» будет затягиваться до тех пор, пока исходная клетка не разделится окончательно на две новые.
В целом описанные выше системы могут показаться очень сложными и запутанными. Однако, если в их устройстве разбираться последовательно, компонент за компонентом, они оказываются очень простыми. У каждого белка своя простая задача. Способность системы выполнять сложные задачи, такие как обнаружение и разделение сестринских хромосом, независимо от их исходного положения в клетке, обеспечивается взаимодействием простых компонентов системы, а не их усложнением. В частности, она связана с тем, что существует обратная связь между поведением компонентов системы и получаемой ими информации о состоянии системы как целого, о том, на какой стадии того или иного процесса она находится в данный момент (например, о том, все ли хромосомы заняли нужное положение). Использование простых компонентов, увязанных в единое целое за счет системы тесных обратных связей, вообще характерно для живой материи, я углубился в детали именно затем, чтобы показать, как системы «глупых» биологических молекул могут организоваться для решения серьезных задач, иначе говоря, чтобы показать, как из простого возникает сложное.
Системы, которые запускали первое деление клетки, продолжают работать в клетках эмбриона, и теперь мы больше не будем обсуждать их в таких деталях. Это вообще типично для биологии: если механизм работает, его можно использовать многократно, иногда с незначительными поправками, в процессе всего эмбрионального развития. Почти сразу после того, как первое деление клетки завершено, каждая из двух образовавшихся клеток начинает копировать хромосомы и делиться. Получается эмбрион из четырех клеток. Синхронные клеточные деления продолжаются некоторое время, но рано или поздно синхронность деления разных клеток теряется, и со стадии примерно шестнадцати клеток число клеток эмбриона начинает отклоняться от значения «два в степени n». Как правило, клетки раннего эмбриона, получившиеся в ходе дробления, расположены довольно свободно относительно друг друга. Иногда, примерно один раз на тысячу двести случаев, клетки распадаются на два отдельных комка. Из каждого комка получится отдельный эмбрион, у которого будет собственная плацента и плодные оболочки. Это один из трех вариантов рождения однояйцевых близнецов, и на него приходится около трети всех случаев. Тот факт, что эмбрион может разделиться и дать начало двум младенцам, говорит нам об очень важной особенности развития: все клетки должны быть способны создать любую часть тела, и нет одной специальной клетки, которая отвечает, например, за создание головы или детерминирована как клетка головы. Если бы клетки с самого начала были разными, если бы одна или несколько клеток были детерминированы к формированию конкретной части тела или если бы одна клетка отвечала за весь процесс, то разделение эмбриона привело бы к тому, что по меньшей мере у одной из половинок не оказалось бы жизненно важных клеток и ее развитие было бы обречено на провал. Тем не менее тысячи живущих в каждой стране однояйцевых близнецов являются красноречивым свидетельством равных возможностей клеток на ранних этапах дробления.
Итак, эмбрион достигает стадии примерно шестнадцати клеток. Это значит, что у него уже достаточно клеток, чтобы изменять форму тела и чтобы начать клеточную дифференцировку, то есть сделать клетки разными. Прелюдия окончена, и начинается основная работа по эмбриональному развитию.
Отрывок из книги Джейми Дейвиса "Онтогенез. От клетки до человека"
2060
2016.10.13 14:36:32
