Как клетка чинит свою ДНК?

Защита генов от мутаций — задача, которую клеткам приходится решать буквально в каждый миг своего существования. у всех живых организмов есть несколько способов отремонтировать повреждённую ДНК, и как раз за расшифровку этих механизмов в нынешнем году присуждена Нобелевская премия по химии. её лауреатами стали Томас Линдаль, Пол Модрич и Азиз Санджар.
Наша наследственная информация, да и не только наша, а вообще любого живого организма на Земле, хранится в молекулах ДНК, зашифрованная с помощью четырёх буквенного генетического кода. Чтобы информация «заработала», нужно с её помощью создать белковую молекулу. Что при этом происходит?
Во-первых, участок ДНК, в котором содержатся сведения о структуре того или иного белка, копируется в молекулу РНК, — информация переписывается с одного молекулярного носителя на другой, происходит транскрипция. Затем на РНК синтезируется собственно белок, полипептидная молекула, — генетическая «инструкция по сборке» воплощается в конкретном «изделии»; происходит трансляция. Но одними лишь белковыми инструкциями информация в ДНК не исчерпывается: разных белков на разных этапах жизни организма требуется разное количество, поэтому в любом геноме есть огромная масса регуляторных «параграфов», которые позволяют очень точно регулировать количество синтезируемого белка в соответствии с обстоятельствами. Наконец, копию всех генов нужно передать потомству, то есть возникает задача удвоить весь свой геном, или, иными словами, совершить репликацию ДНК.
И при синтезе белка, и при репликации ДНК важно, чтобы никаких ошибок в генетическом тексте не появлялось. Замены или потери «букв» в последовательностях генов не всегда приводят к фатальным результатам, однако вероятность такого исхода довольно велика: подобные погрешности-мутации могут либо вообще сделать белок неактивным, либо изменить функцию белка так, что он принесёт больше вреда, чем пользы.
Если же мутация появится в момент синтеза новой ДНК, то вполне возможно, что в результате сломается настолько важная молекула, что потомство просто не сможет появиться на свет. Наконец, изменения в нуклеотидах могут сделать невозможной репликацию или транскрипцию. Чтобы защитить себя от таких неприятностей, живым организмам уже на самых первых этапах эволюции пришлось создать у себя специальные молекулярные механизмы, которые следили бы за состоянием генома и вовремя исправляли бы возникавшие в нём ошибки, то есть, иначе говоря, выполняли бы репарацию, или ремонт, ДНК.
Вместе с репликацией, транскрипцией и трансляцией репарация стала одним из фундаментальных молекулярных процессов, присущих всему живому, и именно за расшифровку механизмов репарации Нобелевский комитет присудил в нынешнем году премию по химии.
Для того, чтобы понять суть репарации, нам нужно хотя бы бегло вспомнить, как устроена ДНК. Мы знаем, что она представляет собой двойную спираль — две цепи обвиты вокруг друг друга. Каждая цепь состоит из огромного количества нуклеотидов. Что такое нуклеотид? Это трёхчастная молекула, состоящая из остатка фосфорной кислоты, соединённого химической связью с сахаром дезоксирибозой, к которой в свою очередь присоединено азотистое основание. Цепочка же получается потому, что фосфорная кислота сцепляется с ещё одной рибозой, от другого нуклеотида, и таким образом два основания становятся соседями. В каждой цепочке ДНК можно выделить так называемый сахарофосфатный остов, на котором, как на заборе, сидят азотистые основания. Именно они и являются теми самыми генетическими «буквами», числом четыре: A — основание аденин, T — основание тимин, G — основание гуанин, C — основание цитозин. (У другой нуклеиновой кислоты, РНК, есть свои отличия в химическом и структурном строении, в которые мы сейчас углубляться не будем.) В чередовании A, T, G и C как раз и заложена информация о структуре белка: стоящие подряд три основания формируют триплет, или кодон, и соответствуют какой-либо аминокислоте (скажем, TTT кодируют аминокислоту фенилаланин, а CTG — аминокислоту лейцин).
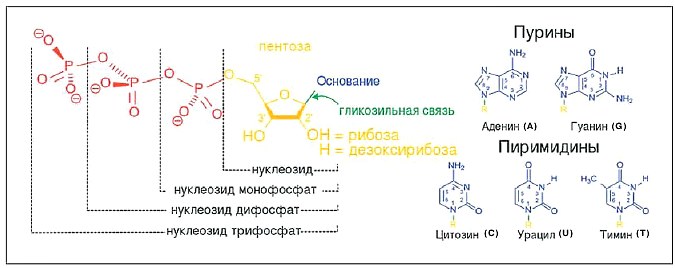
Химическое строение нуклеотидов. ДНК и РНК сложены из нуклеозид монофосфатов, причём каждый фосфат (остаток фосфорной кислоты) соединяется ещё и с рибозой следующего, соседнего нуклеотида. У РНК и ДНК сахар рибоза отличается наличием или отсутствием одного атома кислорода. Синим цветом показаны азотистые основания, которые делятся на две группы: пурины (A, G) и пиримидины (C, U, T). В состав РНК вместо тимина входит урацил.
Химическое строение нуклеотидов. ДНК и РНК сложены из нуклеозид монофосфатов, причём каждый фосфат (остаток фосфорной кислоты) соединяется ещё и с рибозой следующего, соседнего нуклеотида. У РНК и ДНК сахар рибоза отличается наличием или отсутствием одного атома кислорода. Синим цветом показаны азотистые основания, которые делятся на две группы: пурины (A, G) и пиримидины (C, U, T). В состав РНК вместо тимина входит урацил.
Кроме того, A, T, G и C могут образовывать водородные связи друг с другом. Благодаря спариванию нуклеотидов образуется двойная спираль: основания одной цепочки взаимодействуют с основаниями другой цепочки. Взаимодействуют не абы как, а в соответствии со строгим правилом: аденин — с тимином, гуанин — с цитозином. Комплементарное спаривание нуклеотидов поддерживает правильную структуру всей молекулы. Понятно, что, если ген зашифрован в одной цепи, то в другой цепи порядок нуклеотидов будет как бы зеркальным по смыслу, — это называют смысловой и антисмысловой последовательностями.
Какие повреждения могут случиться с ДНК? Например, у неё могут разорваться цепи — или одна, или сразу обе. В клетках есть специальные ферменты, которые ликвидируют разрыв, сшивая оба конца обратно. (Такие белки тоже относятся к репарирующим системам, хотя разрыв не самый сложный, не самый хитрый случай.) Но может быть и
так, что никакого разрыва нет, просто азотистое основание вдруг подверглось химической модификации, и теперь оно совсем не то, каким должно быть. В подобных случаях в клетках существует сразу несколько ремонтных систем.
Дефекты в ДНК могут появиться под действием жёсткого ионизирующего излучения или ультрафиолета. Ещё в 20-е годы прошлого века выдающийся американский генетик Герман Мёллер обратил внимание на то, что бактерии после обработки рентгеновскими лучами перестают расти и гибнут. (Впоследствии Мёллер стал лауреатом Нобелевской премии по медицине за открытие появления мутаций под влиянием рентгеновского излучения.)
Такие же результаты вскоре получили и с УФ-светом. Затем в 1940-е годы микробиолог Альберт Кельнер обнаруживает, что облучённые ультрафиолетом бактерии можно реанимировать, если... просто посветить на них видимым светом. Феномен назвали фотореактивацией, но что за механизм лежит в его основе, никто не знал. В 1944 году благодаря экспериментам Освальда Эвери, Колина Маклеода и Маклина Маккарти стало ясно, что за наследственность в клетке отвечает ДНК (хотя, как именно у неё получается хранить наследственную информацию, не было понятно до 1953 года, когда Уотсон и Крик опубликовали статью с описанием структуры ДНК). В 1950-е годы стало ясно, что ультрафиолет вредит именно ДНК и что у бактерий и дрожжей есть некий фермент, который на свету ликвидирует нанесённый ущерб.
Вскоре к «световой» репарации, то есть фотореактивации, добавилась «темновая», которая шла хоть на свету, хоть в темноте. Заодно удалось понять, чем именно плох ультрафиолет: эксперименты Джейн и Ричарда Сетлоу показали, что УФ-излучение сшивает рядом стоящие в одной цепочке ДНК основания тимины (генетическая «буква» T) в димеры. Тиминовые димеры мешают молекулярным машинам работать с ДНК, однако вскоре исследователи обнаружили, что бактерии (все подобные эксперименты ставили поначалу на бактериях либо на дрожжах) избавляются от модифицированных оснований, попросту вырезая их из ДНК, и что эта процедура не связана с репликацией. Стало ясно, что у клеток есть особый, независимый от других молекулярных процессов механизм, названный эксцизионной (от англ. excision — удаление, ликвидация, вычёркивание) репарацией нуклеотидов, ЭРН или NER (nucleotide excision repair).
Оставалось самое сложное — понять, как именно такая репарация работает.
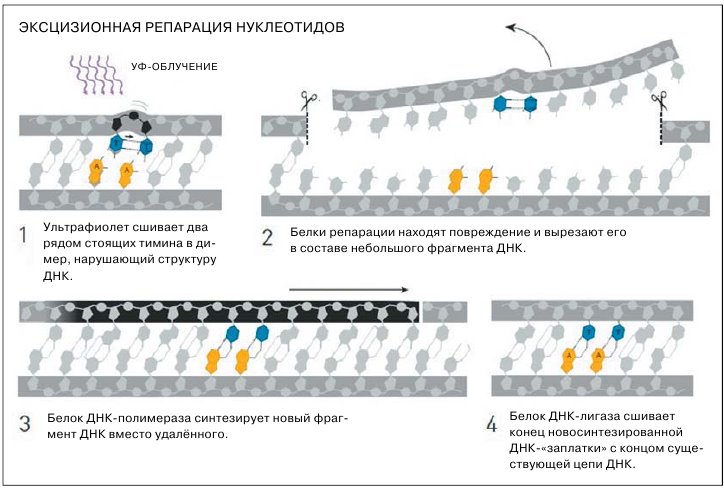
Эксцизионная репарация нуклеотидов включается при повреждениях ДНК, случившихся, например, из-за УФ-облучения или под воздействием канцерогенных веществ табачного дыма
Тут пришла пора вспомнить про первого из нынешних лауреатов, Азиза Санджара (Aziz Sancar). Он родился в Турции в 1946 году, окончил медицинский факультет Стамбульского университета, после чего перебрался в США, где занялся биохимией. В Техасском университете в Далласе Санджар включился в исследования фотореактивации и ему даже удалось найти ген фермента, который отвечает за «световую» репарацию, и получить первые результаты по характеристике этого белка, который был назван фотолиазой. Однако новые данные в то время никого не вдохновили и Санджару пришлось на время забыть про фотолиазу.
Перебравшись сначала в Йельский университет, а потом в университет Северной Каролины в Чапел-Хилл, он занялся механизмом эксцизионной репарации нуклеотидов. К тому времени уже были определены гены, кодирующие для неё белки, uvrA, uvrB и uvrC, но для того, чтобы выяснить детали процесса, нужно было определить, что за белки соответствуют этим генам, а подобная задача в те времена (конец 1970-х — первая половина 1980-х) была ещё весьма непростой.
И вот Санджар придумывает метод, позволяющий выловить нужные молекулы из массы белков, плавающих в цитоплазме бактерии. Он вставляет ген в плазмиду — особую молекулу ДНК, которую можно ввести в клетку, где плазмида будет существовать вместе с бактериальной хромосомой, причём она будет независимо от хромосомы удваиваться (реплицироваться) и на ней будут синтезироваться РНК для белкового синтеза. Фокус же в том, что такую плазмиду вводили в клетки, в которых хромосома была инактивирована; вдобавок в питательную среду, в которой жили бактерии, вносили радиоактивную метку, включавшуюся в белковые молекулы при их синтезе. В результате получалось, что весь новосинтезированный белок можно было детектировать по радиоактивности и весь он происходил с плазмиды — хромосома-то молчала. Вставляя в плазмиду uvrA, uvrB и uvrC, Санджару и его коллегам удалось определить соответствующие белки, после чего уже стало понятно, как их можно очистить в количестве, достаточном для последующих опытов.
В 1983 году выходит статья с описанием механизма NER, который впоследствии ещё несколько раз уточнялся. Сейчас мы знаем, что всё начинается с белкового комплекса, состоящего из двух экземпляров UvrA и одного UvrB: они сканируют ДНК на предмет повреждений, пока UvrA что-нибудь такое не найдёт. Тогда UvrB расплетает в проблемном месте двойную спираль ДНК, чтобы отделить «здоровую» цепь от повреждённой. На место UvrA приходит UvrC, и вот UvrC вместе с UvrB выполняют главное действие: вырезают из повреждённой цепи участок с неправильными нуклеотидами. Причём участок этот вырезается с запасом — дефектные нуклеотиды находятся примерно в центре фрагмента длиной в 12—13 «букв». Ещё один белок, UvrD, помогает удалить вырезанный фрагмент, после чего в дело вступает фермент ДНК-полимераза, — она синтезирует «заплатку» к дыре в повреждённой цепи по шаблону той цепи, которая осталась целой, руководствуясь принципом комплементарности нуклеотидов. Завершают всё другие ферменты, пришивающие «заплатку» к её месту в ДНК.
Поначалу из повреждений ДНК, которые могли быть причинены УФ-излучением, были известны только тиминовые димеры, но потом оказалось, что таких дефектов существует довольно много. Все они распознаются аппаратом эксцизионной репарации нуклеотидов. У высших животных и у человека она тоже работает, с теми же стадиями: узнать — вырезать — залатать. Правда, с течением эволюции увеличилось число молекул-«исполнителей», отвечающих за распознавание повреждения и разрезание ДНК: у бактерий их всего три, а в человеческих клетках под те же задачи выделено целых 15 различных белков.
Если мутации коснутся самих белков эксцизионной репарации, то это может привести к пигментной ксеродерме, — так называют наследственное заболевание кожи, проявляющееся повышенной чувствительностью к ультра-фиолетовому облучению; в дальнейшем всё может закончиться раком. Изучение ксеродермных мутаций помогло в своё время установить, какие белки отвечают за репарацию в человеческих клетках.
В этом большую роль сыграли исследования Джеймса Кливера, Ричарда Вуда и самого Санджара. Он, между прочим, после того, как раскрыл механизм «теневой» репарации, и параллельно с дальнейшими её исследованиями, нашёл время вернуться к своей старой теме, связанной с фотореактивацией. В работах, опубликованных во второй половине 80-х годов XX века, Азиз Санджар описал действие фотолиазы, как она, улавливая энергию света, попросту разделяет сшитые ультрафиолетом димеры нуклеотидов. Так что, как видим, на его счёт можно записать целых два механизма репарации.
Фотореактивация есть у бактерий, архей, дрожжей, насекомых, но у человека, как и у многих высших эукариот, ничего такого нет. Пока что мы говорили о нуклеотидных мутациях, случившихся в из-за внешнего воздействия — ультрафиолетового или ионизирующего излучения. До поры до времени биологи думали, что в отсутствие таких жёстких факторов ДНК вполне стабильна. Однако уже в 1970-е годы стало понятно, что её стабильность заметно преувеличена: эксперименты шведского биохимика Томаса Линдаля (Tomas Lindahl), второго нынешнего лауреата премии по химии, показали, что и в обычных физиологических условиях азотистые основания в ДНК теряют аминогруппы, претерпевают окисление и безо всякой помощи ферментов приобретают метильные группы. Одно из самых характерных изменений происходит с цитозином (генетическая «буква» C): он спонтанно теряет аминогруппу, после чего превращается в основание урацил (U), который является аналогом тимина. Цитозин, как мы помним, комплементарно спаривается с гуанином, но у урацила, как и у тимина, в напарниках аденин. После превращения C в U в этом месте ДНК возникнет неправильное нуклеотидное спаривание, последовательность кода окажется нарушенной. При синтезе новой цепи ДНК во время репликации или при синтезе РНК во время транскрипции ошибка либо перейдёт в следующее поколение, либо испортит белок. Похожие проблемы возникают и при других модификациях азотистых оснований.
Линдаль заметил, что урацил появляется в ДНК достаточно часто, однако никакой катастрофы не происходит. Иными словами, в клетке должен быть механизм, который такие ошибки исправляет. Линдалю удалось в буквальном смысле в одиночку найти бактериальный фермент урацил-ДНК-гликозилазу, которая точечно вырезала неправильное основание из цепи ДНК; вскоре к ней добавилась ещё одна гликозилаза, выщеплявшая метилированный аденин. Сейчас таких ферментов открыто уже много, все они входят в механизм эксцизионной репарации удалением повреждённых оснований, ЭРО или BER (base excision repair).
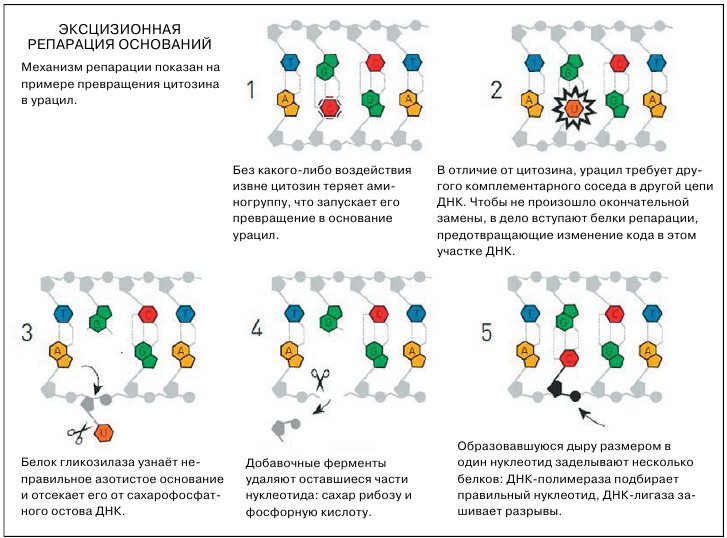
Эксцизионная репарация оснований включается при спонтанных химических модификациях азотистых оснований.
Название похоже на эксцизионную репарацию нуклеотидов, однако у них есть важное отличие: если при эксцизионной репарации нуклеотидов из ДНК полностью выщепляется нуклеотид, и не один, а целая шеренга, то здесь гликозилазы отделяют собственно неправильное, модифицированное азотистое основание, саму «букву»; сахарофосфатный остов же остаётся нетронутым. Но лишь на время — вслед за гликозилазами к месту дефекта приходят другие ферменты из группы эндонуклеаз, которые специализируются именно на таких точках, где есть сахар, есть фосфорная кислота, но нет азотистого основания (некоторые из них были описаны ещё за два года до того, как Линдаль открыл свои гликозилазы). Эти эндонуклеазы вырезают оставшиеся компоненты нуклеотида, то есть фосфорную кислоту и присоединённый к ней сахар. Затем появляется ДНК-полимераза, которая подбирает для образовавшейся дыры правильный нуклеотид, согласно принципу комплементарности, и специальные ферменты вшивают его на место.
Как и в случае с эксцизионной репарацией нуклеотидов, все компоненты BER в конце концов собрали in vitro, в реакционной смеси, чтобы подтвердить, что всё работает так, как описано. И, как обычно, сначала всю систему исследовали на бактериях, а потом перешли на человеческие клетки. Сейчас известно огромное количество различных неправильных модификаций оснований (одних только окислительных дефектов насчитывается более сотни), возникают они не только спонтанно, «сами по себе», но и под действием внешних факторов, вроде ионизирующего излучения. Большую их часть ликвидирует именно аппарат BER, в котором главную роль играютгликозилазы, специализирующиеся на узнавании тех или иных неправильных азотистых оснований.
Пол Модрич, третий нобелевский лауреат, раскрыл ещё один механизм репрации, непосредственно связанный с репликацией ДНК. Молекулярные машины, которые занимаются репликацией, выполняют её не со стопроцентной точностью, то есть в синтезируемые цепочки всё-таки вкрадываются кое-какие ошибки, появляются неправильно спаренные основания. У репликативного аппарата есть, скажем так, встроенная способность исправлять собственные ошибки, но всё равно какие-то из них остаются. И вот тогда в дело вступает репарация ошибочно спаренных нуклеотидов, или мисмэтчрепарация (от англ. mismatch — плохое сочетание, несоответствие). Здесь речь идёт не о модифицированных основаниях, как в предыдущем случае, а о вполне обычных, стандартных A, G, C и T, которых вдруг поставили напротив неправильного соседа.
Впервые сам феномен исправления ДНК с плохими парами нуклеотидов заметили ещё в 1960-е годы, а в 1976 году американские исследователи Роберт Вагнер и Мэтью Мезельсон сделали важное наблюдение: оказалось, что исправления происходят только в одной из двух цепочек ДНК. Это кажется вполне логичным: ведь неправильные нуклеотиды появляются только в новосинтезированной цепочке, и старую, то есть ту, которая служит шаблоном, исправлять совсем не надо. Встал вопрос, как ремонтный аппарат отличает новую цепь ДНК от старой, и следом возникло подозрение, что здесь играют роль метильные метки на азотистых основаниях.
Пол Модрич начинал с исследований ферментов ДНК-метилтрансфераз (или ДНК-метилаз) у бактерий. Их функция состоит в том, чтобы метильными группами метить основания в бактериальной ДНК: если потом в клетку проникнет вирус, то его ДНК будет уничтожена как не имеющая защитных метильных меток. Ферменты ставят метилы на обе цепи ДНК, однако при репликации новая цепь какое-то время остаётся без метильных групп, и вот тут её как раз могут проверить машины мисмэтч-репарации.
К 1983 году Модрич, который к этому времени работал с Мезельсоном, сумел прямо доказать, что именно метилирование управляет ходом репарации. К тому времени генетики уже обнаружили гены, необходимые для этого процесса, названные mutH, mutL, mutS и uvrD. Теперь нужно было расшифровать сам механизм, что и было сделано за несколько лет работы: в 1989 году Пол Модрич публикует статью, в которой описывает работу белков мисмэтч-репарации в реакционной смеси, in vitro.
Один из них, MutS, узнаёт неправильное соответствие нуклеотидов в ДНК, другой, MutH, определяет место, где на одной цепи есть метильная группа, а на другой — нет. Расстояние между полуметилированным участком и тем местом, где случилось неправильное спаривание нуклеотидов, может достигать 1000 «букв». Третий главный белок, MutL, передаёт сигнал от MutH к MutS: MutH как бы говорит, какая из двух цепей новая, а MutS делает в новой цепочке надрез рядом с неправильным нуклеотидом. Затем подключается UvrD, который представляет собой хеликазу, то есть фермент-расплетатель: он разворачивает двойную спираль, помогая вытащить фрагмент цепи с дефектом.
Дополнительные белки помогают его удалить, а образовавшуюся дыру — залатать, используя как шаблон для латки всё ту же старую, родительскую, цепь ДНК (на этом этапе мисмэтч-репарация напоминает санджаровскую репарацию нуклеотидов, где тоже из ДНК удаляется целый фрагмент с повреждением). Впоследствии оказалось, что такой механизм весьма консервативен, он относительно мало менялся в ходе эволюции. В 2004 году Пол Модрич сумел собрать из очищенных компонентов человеческую систему мисмэтч-репарации, подобно тому, как в 1980-е он собрал бактериальную систему. Правда, пока непонятно, как у эукариот репаративная машина отличает правильную новую цепь ДНК от неправильной старой. Метилирование тут уже явно ни при чём, у эукариот оно нужно для других вещей.
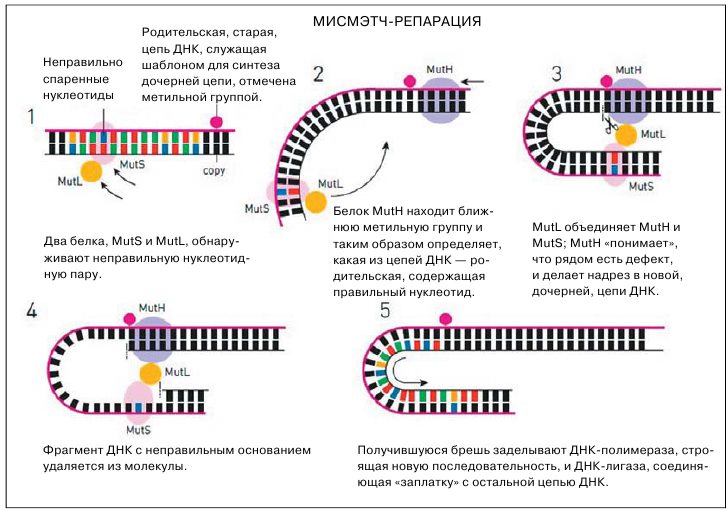
Мисмэтч-репарация следит за ДНК в момент её репликации, когда клетка собирается делиться и удваивает свой геном. Во время репликации на новой цепи ДНК порой появляются нуклеотиды, не подчиняющиеся принципу комплементарного спаривания, и задача мисмэтч-репарации — такие ошибки исправлять.
Среди систем репарации ДНК есть и другие разновидности и модификации, не исключено, что в будущем к ним добавится ещё что-нибудь. Однако три вышеописанных механизма можно назвать основными, присутствующими у большинства живых организмов, и наиболее хитроумными. Если говорить о «народнохозяйственной пользе», то здесь всё просто: ДНК в наших клетках ломается постоянно, и, если её вовремя не ремонтировать, наш геном погрузится в хаос, со всеми вытекающими последствиями.
Чтобы долго про это не говорить, приведём вот такую статистику: считается, что от 80 до 90% всех раковых заболеваний связаны с неправильной работой ДНК-репарирующих систем. В качестве конкретного примера можно ещё раз вспомнить пигментную ксеродерму, о которой шла речь выше, или наследственный неполипозный колоректальный рак, который развивается вследствие мутаций в генах мисмэтч-репарации. С фундаментальной же точки зрения понимание ДНК-репаративных механизмов важно для самой главной биологической науки — эволюционного учения. Дело в том, что мутации совсем не всегда плохо, они часто оказываются хорошими и полезными. Именно мутации служат «сырьём» для эволюции, именно благодаря им появляются более приспособленные особи, поддерживающие существование вида. Однако если бы репарирующие машины работали с абсолютной точностью и не пропускали ни единого дефекта, никаких мутаций и быть бы не могло.
Действительно, точность репарации, как и любых других молекулярных процессов, не стопроцентна, что, очевидно, может быть важным условием эволюционного развития. Когда речь идет об исследованиях таких фундаментальных процессов, то известия о Нобелевской премии воспринимаются с некоторым удивлением — кажется, что за это уже давно всё всем дали. И здесь стоит отдельно сказать о некоторых важных вещах. Во-первых, по ходу рассказа у нас наряду с «нобелевскими фамилиями» то и дело возникал кто-то ещё. Если бы мы задались целью упомянуть всех, чей вклад в расшифровку механизмов репарации был относительно велик, то статья увеличилась бы вдвое. Таков мир молекулярной биологии: всякий результат здесь есть плод усилий огромной массы людей. Тем не менее в каждом открытии можно найти человека, чьи концепции, модели, методы работы оказались решающими. Во-вторых, мы постоянно подчёркивали слово «расшифровка». Сама по себе мысль, что у клеток должен быть механизм ремонта ДНК, довольно очевидна. Совсем другое дело — описание механизмов молекулярных процессов, детализация, определение этапов и пр.
Именно на подтверждение первоначальной догадки во всех трёх случаях ушли годы исследовательской работы, а потом должны были пройти ещё годы для того, чтобы стало окончательно ясно, что всё именно так, как мы выяснили. И потому в формулировке Нобелевского комитета говорится именно о «механизмах репарации ДНК».
Наконец, последнее. Мы почти ничего не говорили о том, к каким странам принадлежат новые лауреаты, в каком университете они выполнили свои эпохальные работы и т. д. Однако современная наука такова, что вопрос о «национальной принадлежности» того или иного открытия просто бессмыслен. Наши герои — прямое тому доказательство: Азиз Санджар имеет двойное гражданство: США и Турции; швед Томас Линдаль, поработав в США, вернулся в Европу, где в разное время работал в Швеции и Великобритании; Пол Модрич хотя и родился, и учился, и работал в США, но при этом по ходу своей карьеры сменил целый ряд университетов, от Стэнфордского до Университета Дьюка. Вывод же отсюда прост и очевиден — наукой стоит заниматься там, где вы сможете найти ответы на свои вопросы и где вы сможете получить результаты, которые принесут пользу всем людям.
Кирилл Стасевич
Источник: Журнал «Науки и Жизнь», № 11, 2015 г.
4317
2015.11.29 11:47:34