Ранние этапы адаптации предсказуемы, поздние — случайны
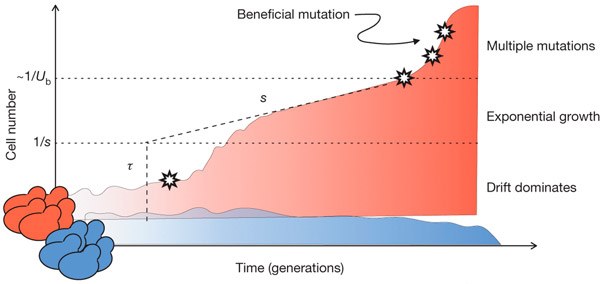
Рис. 1 Типичная судьба клонов бесполых организмов в ходе адаптации. Синим цветом показана динамика численности клона дрожжей, в котором не возникло полезных мутаций. Такой «нейтральный» клон, скорее всего, не достигнет высокой численности и будет вытеснен более успешными конкурентами. Красным цветом показана судьба клона, в котором возникла полезная мутация (показана звездочкой). Такой клон тоже может вымереть, пока его численность мала, за счет случайных колебаний численности (дрейфа). Чтобы отбор начал поддерживать носителей полезной мутации, клон должен достичь пороговой численности, которая для мутации с полезным эффектом s примерно равна 1/s. Например, мутация, повышающая приспособленность на 1% (s = 0,01), начнет поддерживаться отбором при численности мутантов примерно в 100 особей. После прохождения этого порога клон, несущий полезную мутацию, вступает в фазу экспоненциального роста численности, причем по параметрам этого роста можно оценить s и τ (время возникновения мутации). Когда численность клона приблизится к 1/Ub, где Ub — частота возникновения полезных мутаций, в нем с большой вероятностью начнут появляться дополнительные полезные мутации. Шкала по вертикальной оси логарифмическая. Рисунок из обсуждаемой статьи в Nature
Новая методика индивидуального генетического «штрих-кодирования» позволила американским ученым в деталях изучить процесс накопления полезных мутаций в большой бесполой популяции дрожжей в ходе адаптации к новой среде. Как выяснилось, на начальных этапах общий рост приспособленности популяции идет в основном за счет высоковероятных мутаций со слабым положительным эффектом, которые возникают независимо у множества особей. На этой стадии процесс адаптации во многом предсказуем. В дальнейшем роль случайности возрастает, потому что на первый план выходят маловероятные мутации с сильным полезным эффектом.
Задача количественного описания эволюции больших бесполых популяций имеет не только теоретическое, но и практическое значение. Большие, быстро эволюционирующие популяции одноклеточных паразитов, будь то болезнетворные бактерии или раковые клетки, ежегодно уносят жизни примерно 20 миллионов людей (около 30% всех смертей на планете). Чтобы сдерживать эту угрозу, важно знать, какие факторы влияют на рост приспособленности клонально размножающихся одноклеточных и как можно этот рост затормозить.
В популяциях, насчитывающих миллионы особей, в каждом поколении возникает множество новых мутаций, в том числе — «полезных», то есть ускоряющих размножение. Все они вносят вклад в общую приспособленность популяции, от которой зависит скорость ее роста. Но как уследить за тысячами мутаций, происходящими у миллионов особей? Секвенировать целиком миллионы геномов — неподъемная задача даже при современном уровне развития биотехнологий. Если же применять выборочное полногеномное секвенирование, то в поле зрения исследователей попадут только мутации, достигшие высокой численности (частоты встречаемости). Это может существенно исказить картину. Ведь большинство возникающих в бесполой популяции полезных мутаций, возможно, никогда не становятся массовыми, но свой вклад в общую приспособленность они, тем не менее, вносят.
Альтернативный подход состоит в том, чтобы пометить отдельные клоны (клетки, произошедшие от одной и той же родительской клетки) наследуемой меткой, а потом следить, как меняется численность клона. Если она вдруг начала экспоненциально расти (см. Exponential growth), в то время как общая численность популяции остается постоянной, значит, у одного из представителей клона возникла полезная мутация. При этом скорость экспоненциального роста является мерой полезности мутации. Например, если рост численности клона описывается уравнением N = N0·(1 + 0,05)^t, где время (t) измеряется в поколениях, то мутация повысила приспособленность на 5% (полезность мутации s = 0,05). Пример использования такого подхода описан в новости В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011. Но там речь шла только о двух помеченных клонах, что позволяло зарегистрировать лишь одну (первую) полезную мутацию, возникшую в популяции.
Новая статья американских биологов, опубликованная на сайте журнала Nature, демонстрирует важный прорыв в развитии методов наблюдения за эволюцией многомиллионных бесполых популяций. Авторы работали с популяциями дрожжей, в которых были помечены индивидуальными генетическими метками по 500 000 индивидуальных клонов. Для этого была изготовлена большая коллекция плазмид, содержащих случайные двадцатинуклеотидные последовательности (генетический «штрих-код»). Эти плазмиды внедрялись в дрожжевые клетки, геномы которых были предварительно модифицированы таким образом, чтобы плазмиды встраивались в строго определенное место генома при помощи Cre-рекомбиназы (см. Cre recombinase). Кроме того, у подопытных дрожжей была удалена и перенесена в плазмиды часть гена URA3. Это позволило исследователям отобрать те дрожжевые клетки, у которых встраивание плазмиды прошло успешно (только у этих клеток ген URA3 нормально работал, и поэтому они могли расти в среде, не содержащей урацила или уридина (см. Uridine)).
В итоге авторы получили две подопытные популяции дрожжей численностью по 10^8 особей каждая, происходящие от одной предковой клетки. В обеих популяциях каждая клетка принадлежала к одному из полумиллиона помеченных клонов. Эти популяции затем в течение 168 поколений адаптировались к «голодной» среде, где размножение ограничивалось количеством глюкозы.
Численность каждого клона отслеживалась путем массового секвенирования небольшого фрагмента генома, содержащего штрих-код. Секвенировать приходилось лишь 0,002% генома, что позволило резко увеличить «частотное разрешение» метода по сравнению с полногеномным секвенированием. В поле зрения исследователей попали даже те мутации, частота встречаемости которых в популяции никогда не превышала 10^–5, тогда как секвенирование полных геномов позволило бы отследить лишь клоны с относительной численностью от 10^–2 и выше. В результате вместо 25 000 зарегистрированных мутаций исследователям удалось бы обнаружить лишь около 15.
Впрочем, даже зная численность каждого клона в разные моменты времени, определить, в каком из них возникла полезная мутация — не такая простая задача (рис. 1). Каждая мутация возникает сначала у одной особи. Пока число потомков удачного мутанта невелико, динамика их численности определяется не столько приспособленностью (отбором), сколько случайными колебаниями (дрейфом). Большая часть вновь возникающих полезных мутаций теряется из-за дрейфа: потомки удачного мутанта просто не успевают достичь такой численности, при которой отбор «заметит» их полезную мутацию и начнет ее поддерживать. Мутация становится заметна для отбора (и выходит из-под власти дрейфа) лишь по достижении численности, сопоставимой с 1/s. Например, мутация, повышающая приспособленность на 1%, становится заметна для отбора при числе мутантов около 100. Мутантному клону должно повезти, чтобы его численность за счет дрейфа доросла до этого порога — лишь тогда за дело возьмется отбор, а мутацию можно будет считать «установившейся» (established). Такая мутация, если она полезна, уже не потеряется из-за случайного дрейфа.
После того как мутация «установилась», численность клона будет экспоненциально расти. По скорости роста можно оценить полезность мутации (s), а экстраполируя тренд в прошлое, можно примерно определить время возникновения мутации (τ). Впрочем, этот удобный для количественного анализа период в истории клона будет продолжаться лишь до тех пор, пока в геноме, уже содержащем одну полезную мутацию, не возникнет вторая. После этого все расчеты оказываются резко затруднены и картина смазывается. Именно поэтому рассматриваемая методика позволяет анализировать лишь начальные этапы адаптации — до тех пор, пока вероятность возникновения повторных полезных мутаций остается пренебрежимо малой (а она перестает быть таковой, когда численность клона приближается к 1/Ub, где Ub — вероятность возникновения полезной мутации в расчете на особь за одно поколение).
Авторы использовали множество сложных статистических приемов и компьютерное моделирование, чтобы учесть все эти и многие другие методологические трудности. В итоге они пришли к следующим выводам.
Численность большинства клонов снижалась в ходе эволюционного эксперимента, нередко — до полного вымирания (синие линии на рис. 2). Такая судьба характерна для «нейтральных» клонов, в которых не возникло полезных мутаций. Однако примерно у 5% клонов (25 000 из 500 000) за первую сотню поколений появилась и «установилась» полезная мутация (красные линии). Речь идет о 100 поколениях, а не о всех 168, потому что полезные мутации, появившиеся на поздних этапах эволюционного эксперимента, не могли быть зарегистрированы при помощи применявшихся методик: эти поздние мутации просто не успевали набрать достаточную численность за время, оставшееся до конца эксперимента.
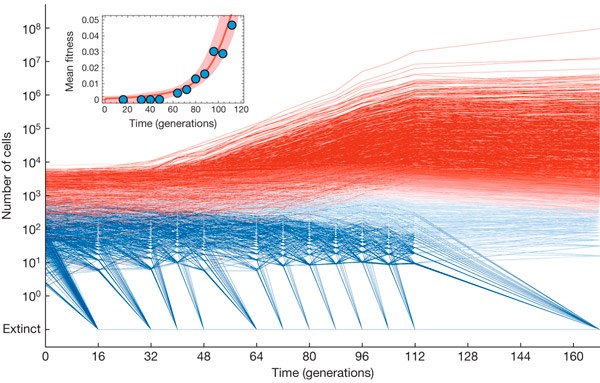
Рис. 2 Динамика численности клонов в подопытной популяции дрожжей. Разными цветами показана оценка вероятности того, что в данном клоне произошла полезная мутация (красные — удачные мутанты, синие — «нейтральные» клоны). Снижение численности многих клонов с полезными мутациями к концу эксперимента объясняется ростом средней приспособленности популяции: многие слабополезные мутации, которые поначалу давали преимущество, начали клониться к упадку, когда в популяции стали преобладать клоны-конкуренты с более высокими s. График на врезке показывает динамику средней приспособленности популяции. Рисунок из обсуждаемой статьи в Nature
По мере размножения клеток, обладающих полезными мутациями, средняя приспособленность популяции росла. Это, в свою очередь, приводило к тому, что «нейтральные» клоны, в которых не было полезных мутаций, а приспособленность осталась на исходном уровне, оказывались во всё более проигрышном положении. Их численность продолжала убывать. Более того, многие «удачливые» клоны, поначалу бодро размножавшиеся благодаря рано возникшей слабополезной мутации, в дальнейшем приходили в упадок и начинали вымирать. Это объясняется тем, что на фоне возросшей приспособленности конкурентов их полезная мутация уже не давала преимущества. Именно поэтому многие красные линии начинают идти вниз в правой части графика на рис. 2. Средняя приспособленность популяции за 100 поколений выросла примерно на 5%. Соответственно, к этому времени все обладатели мутаций с s
Эти результаты говорят о том, что ключевым процессом, определяющим динамику адаптации в большой бесполой популяции является безжалостная конкуренция между клонами — обладателями разных полезных мутаций (клональная интерференция, см. Clonal interference). При этом ранние стадии адаптации определяются в основном высоковероятными и потому многочисленными слабополезными мутациями. Таких мутаций, повышающих приспособленность на 2–5% (0,02 0,1). На этом этапе ход адаптации становится всё менее предсказуемым, поскольку то, в какие моменты и в каком количестве возникнут в популяции эти маловероятные мутации, зависит от случая.
Внимательный читатель может заметить кажущееся противоречие между этими результатами и теми, которые были получены в долгосрочном эволюционном эксперименте Ричарда Ленски (см.: Новые результаты долгосрочного эволюционного эксперимента: приспособленность подопытных бактерий продолжает расти, «Элементы», 23.12.2013). Там сначала закреплялись мутации с сильным положительным эффектом, и лишь затем начинали фиксироваться мутации, ненамного повышавшие приспособленность. На самом деле расхождений тут нет, просто речь идет о разном временном масштабе изучаемых процессов и о разных этапах адаптации (у Ленски — 50 000 поколений, в рассматриваемой работе — 168), а также о разных отслеживаемых параметрах и разной разрешающей способности методов. В работе Ленски отслеживались в основном зафиксировавшиеся, то есть достигшие 100-процентной частоты полезные мутации, а в рассматриваемой работе речь идет лишь об «установившихся» мутациях, подавляющее большинство которых никогда не зафиксируется, а будет вытеснено в ходе клональной интерференции. Те мутации, которые зафиксировались бы в итоге у подопытных дрожжей (если бы эксперимент продлился еще несколько сотен поколений), были бы как раз мутациями с сильным положительным эффектом, как и в эксперименте Ленски. Новое исследование раскрыло тонкие детали начальных этапов адаптации. В частности, оно объяснило, почему многочисленные слабополезные мутации поначалу не имеют шанса зафиксироваться: они будут неизбежно отбраковываться по мере роста общей приспособленности популяции.
Авторы оценили также общую частоту возникновения полезных мутаций у подопытных дрожжей после того, как их поместили в «голодные» условия. Полезная мутация с s > 0,05 возникает в такой ситуации с частотой 10^–6 на клетку за поколение. Приняв во внимание базовую частоту мутирования исходного штамма (4·10^–10 на нуклеотид за поколение), авторы рассчитали ту долю неповторяющихся частей генома, изменения в которой в данных условиях повышают приспособленность на 5 и более процентов. Эта доля оказалась равной 0,04% (5000 нуклеотидов). Это примерно соответствует оценкам, полученным ранее другими авторами. Впрочем, данная цифра сильно зависит от условий, к которым нужно приспосабливаться. Понятно, что если поместить дрожжи в идеальные для них условия, то вероятность того, что случайная мутация окажется полезной, будет крайне низка. Трудно что-то улучшить, когда и так всё прекрасно (см. Принцип Анны Карениной). Соответственно, в плохих условиях эта вероятность будет выше.
Многие теоретики предполагали, что частота возникновения мутаций с тем или иным значением s (μ(s)) должна экспоненциально убывать по мере роста s. Полученные результаты не подтвердили эту гипотезу. Хотя μ действительно снижается с ростом s, это снижение не похоже на монотонную экспоненту. Например, оно резко замедляется в диапазоне s от 0,08 до 0,10, так что график на этом участке почти выходит на плато (рис. 3). Авторы предполагают (и приводят ряд статистических аргументов в пользу своего предположения), что плато соответствует одному конкретному классу мутаций, а именно мутациям, отключающим или выводящим из строя тот или иной ген, работа которого в новых условиях не идет на пользу организму. Известно, что такие мутации нередко оказываются полезными и закрепляются на самых ранних этапах адаптации к новым условиям.
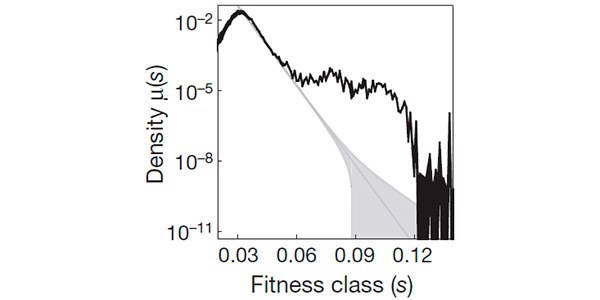
Рис. 3 Зависимость частоты возникновения полезных мутаций μ(s) от их полезности s. По вертикальной оси отложена «плотность» μ(s); реальная частота возникновения мутаций в определенном диапазоне значений s равна площади под соответствующим участком кривой. Например, частота возникновения мутаций с s > 0,05 примерно равна 10^–6. Рисунок из обсуждаемой статьи в Nature
Исследование наглядно показало, что «генетическое штрих-кодирование» позволяет изучать ранние этапы адаптации больших бесполых популяций с очень высоким разрешением, замечая полезные мутации, частота которых не достигает даже сотых долей процента. К важнейшим и относительно неожиданным результатам относится вывод о том, что начальный рост общей приспособленности популяции обусловлен в основном теми полезными мутациями, которым не суждено достичь высоких частот и которые вскоре будут вытеснены. Таких мутаций оказалось неожиданно много.
Недостатком метода является неизбежное снижение разнообразия штрих-кодов в популяции со временем. Из-за этого метод пока пригоден только для анализа самых ранних этапов адаптации. Но это ограничение можно преодолеть, если научиться добавлять в популяцию новые штрих-коды по мере необходимости.
Александр Марков
Источник
1068
2015.03.05 13:01:52